
📖🧬 Qué aporta la cladística a las conductas que no fosilizan
Como psicólogo evolucionista me encuentro muchas veces falta de comprensión, que no de noción, sobre la teoría evolucionista. Sé perfectamente que nuestra disciplina, que se centra en el estudio multidisciplinar de la conducta y cognición de nuestra especie a nivel filogenético, presenta muchos límites. Algunos abordables y otros no. Casi todos estos límites parecen estar radicados en la perspectiva y modelo explicativo que uno sigue. Por eso a veces nos encontramos con just so stories, relatos explicativos que parecen saltos mortales con tirabuzón y con, sencillamente, falta de formación e idea en varios aspectos que considero primordiales a la hora de abordar esta temática, partiendo de modelos teóricos en apariencia inamovibles: la conducta no fosiliza; la relación entre ontogenia, ecología del comportamiento y filogenia es más que clara; la naturaleza cultural de nuestras conductas es el motor que aceleró nuestra evolución como especie y no desprecia a lo biológico, sino que lo justifica y lo explica; y la cladística, que incluye lo que tenemos en común o no con otras especies, es a veces ignorada o malinterpretada.
El primero es un problema más que obvio. La conducta no fosiliza y debemos realizar inferencias continuamente. Desde la paleoneurología a la arqueología cognitiva, desde la traceología a la paleoecología. Se debe hacer hincapié, y no se hace, sobre que estas inferencias deben centrarse en trazar tanto los límites como el potencial de la conducta de las distintas especies, no el determinisimo a la hora de explicar una conducta. Es algo de lo que todos los que hemos empezado en algún momento a indagar en esta temática nos hemos dado cuenta que habíamos hecho alguna vez. Todos somos pecadores. Por ejemplo, si yo digo que la corteza parietal se ha expandido en los seres humanos modernos puede explicar que tuvimos el potencial de manipular objetos con una motricidad cada vez más fina, pero no que la tuvimos únicamente porque se expandió nuestra corteza parietal o que se redujo la cresta sagital por mutaciones en varios genes de la miosina. Condiciones necesarias y que no son excluyentes, pero tampoco son suficientes.
En segundo lugar, el enfoque eco-evo-devo en Psicología Evolucionista intenta resolver algunos problemas explicativos de otros que han tenido un mayor relato y presencia a lo largo de estas últimas décadas, como la escuela de Santa Bárbara, centrada sobre todo en el correlato biológico-conductual, la selección sexual, la modularidad cognitiva y la Era de las Adaptaciones, siendo la más conocida y vilipendiada. Ignora que no tenemos módulos, sino procesos, dominios y sistemas cognitivos, lo que en apariencia puede parecer lo mismo, pero no lo es. No es lo mismo decir que tenemos módulos cognitivos para cocinar o cazar, que se asemeja a la gilipollez de las inteligencias múltiples, a decir que tenemos procesos atencionales, mnésicos, ejecutivos y lingüísticos productos, todos ellos, de la interrelación de sistemas aferentes y eferentes, sensibles a su vez a las leyes del aprendizaje y al entorno sociocultural.
Tampoco tuvimos una era de las adaptaciones con un Gran Salto Adelante en el Paleolítico Superior Europeo. Toda nuestra evolución como especie es una era de las adaptaciones, incluso antes de ser humanos, antes de ser homininos o primates. Es cierta la aceleración cultural hace 45 kya, como lo es la de la Middle Stone Age hace 77 kya, como asu vez lo fue la aparición de los HAM hace entre 320 y 200 kya, la aceleración hace 500-400 kya de los preneandertales y el fuego, etc. Nos gustan los hitos, ponerles nombres y organizar un relato tras ello, sobre todo si eso implica a Europa. Sin embargo, hay que prestarle más atención a la historia evolutiva, a los tiempos y al mosaicismo propio de todo proceso evolutivo. También me produce dentera la obsesión por la selección sexual.
Es obvio que el sexo biológico tiene una importancia clara a la hora de explicar la variabilidad conductual, y fenotípica en general, de cualquier especie pero con nosotros existe un problema doble: no hace hincapié en su relación ni con la selección social o ecología del comportamiento (véase Dunbar) como con la relación entre filogenia y ontogenia. La variabilidad individual, la prevalencia de perfiles en una población y la diversidad de fenotipos quedan fuera del brochazo grueso de estos discursos. Véase por ejemplo la risible pirueta que se han hecho siempre con los postulados de Dart y la caza, ignorando por complejo a la mujer biológica en este aspecto. O hablar de virtue signaling centrándose únicamente en aspectos de competencia intrasexual, ignorando que la virtud es más que relativa (salvo honrosas excepciones) en nuestra especie. Otro ejemplo que no me deja dormir es confundir el adaptacionismo con la selección positiva de rasgos, como puede verse en los fenotipos donde el autismo está presente.
No es lo mismo decir que tal rasgo es una adaptación porque está presente en prácticamente la mayoría de los seres humanos, a que el autismo es otra adaptación porque está presente en un grupo determinado de la población, con su variabilidad propia. Es decir, la selección positiva de rasgos explica mejor la diversidad de estos que forman nuestra variabilidad conductual, así como los subproductos evolutivos, las compensaciones y el ruido en la genética de las poblaciones. No se trata de ser moderno, ni woke, sino de establecer correlatos con un mínimo de sensatez. A su vez, la importancia de nuestra ontogenia, que parece olvidada, no trata única y exclusivamente de dar ese papel cada vez más relevante a la neotenia, altricialidad y a la longevidad de cada uno de nosotros, sino explicar que precisamente por esto, por ser hipersensibles a lo largo de una vida larga, de madurez tardía y dependencia contextual, es lo que nos lleva al cuarto punto que es la selección cultural. Somos animales especialmente sensibles a la cultura, como diría Tomasello. Primero por ser animales que crean nicho y a su vez por ser primates, pues nuestro orden ha encontrado en la cultura el puente adecuado entre entorno social y físico, lo que nos vuelve a llevar a la ecología del comportamiento.
Si observáis al resto de primates veréis que algunos son capaces de crear tradiciones culturales. Es importante el cociente de encefalización para observar conductas sociales complejas, pero necesitas de entorno social complejo (por ejemplo ser animales solitarios no favorece, como pasa con los lémures, y en otros no es importante, como pasa con los orangutanes), poseer una conducta trófica que implique una presión selectiva diferencial, como pasa con los antropoides y su dependencia por la fruta madura y estacional, y, claro, tener de cultura acumulativa. O más bien la capacidad de que la relación entre esa función de emular, imitar, innovar y transmitir nociones culturales de una generación a otra sea otra parte del estudio en Psicología Evolucionista, como pasa con las relaciones genes-cultura. No es lo mismo que un animal sea sensible a la mímesis o al contagio conductual ante la presencia intraespecífica que lo sea al aprendizaje pedagógico, ni que un animal esté determinado prácticamente en todos sus aspectos cognitivos y conductuales por el lenguaje, como pasa con nosotros que tenemos encéfalos especialmente grandes, así como una cognición contaminada por la visión, manos prensiles y una metacognición sociocultural, a que un capuchino Sapajus fabrique o manipule industria lítica para machacar nueces. Ambas presiones, lenguaje e industria lítica, son aspectos culturales pero los caminos evolutivos de ambos taxones han sido completamente distintos, lo que nos lleva al último punto que es la cladística.
Ésta es la rama de la Biología que se centra en las relaciones de los organismos en cuanto a similitudes y diferencias, esos parecidos razonables que a veces no lo son tanto y que otras muchas producen divisiones irreconciliables tanto en la Psicología Comparada como en la Primatología, que en mi opinión deberían ser los pilares más importantes de la Psicología Evolucionista, junto a la Psicología del Desarrollo. Es decir, se trata del análisis de las sistemáticas filogenéticas que observamos en las relaciones de las distintas especies y lo que ha producido que existan dos corrientes principales en nuestros campos de estudio, como es el rupturismo (Povinelli) y el continuismo (de Waal). El primero se centra en qué cambios cuantitativos han producido cambios cualitativos, como las presiones culturales que han conformado un sistema nervioso tan complejo como el nuestro, y el segundo en los rasgos que nos unen a las especies, viendo los cambios como parte de un continuo.
Si bien normalmente es un aspecto central en el estudio de la Morfología de las especies, veo que es especialmente útil también en otros elementos como son los conductuales y cognitivos. El mundo es para los valientes, como diría mi padre. Por ejemplo, es interesante observar una apomorfía (rasgo derivado o novedoso de un grupo monofilético) como es la bipedestación siendo el tipo de desplazamiento principal en los seres humanos, que es derivado a su vez de una sinapormofía (rasgo novedoso compartido por todo un taxón) como lo es la ausencia de cola en Hominidae y que fue novedoso en su momento partiendo de una plesiomorfía (rasgo ancestral) como lo fue la presencia de cola, o vivir y desplazarse a su vez por el dintel de los sistemas boscosos.
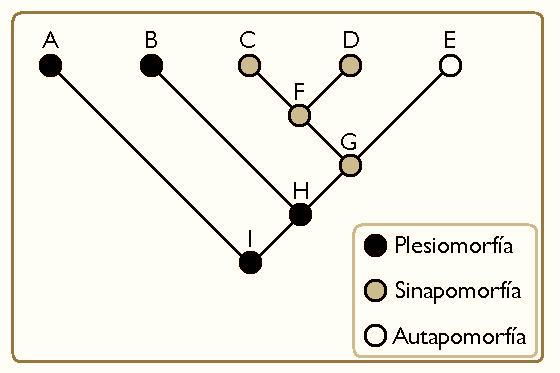
Lo interesante de estos aspectos es implicar rasgos que van más allá de lo morfológico y lo ecológico, yendo a lo puramente conductual. Si separamos lo que tenemos en común de lo que tenemos como propio, no solo nuestra especie sino cualquier otra. Es decir, sería interesante explicar por qué si los Cercopithecini tienen dominancia hemisférica inversa a la nuestra también poseen sistemas de comunicación complejos, como son las vocalizaciones y la semántica; por qué un cuervo de Nueva Caledonia tiene una mayor cognición implicada en el uso instrumental de herramientas que un tití pigmeo (y no me vale que me digáis que estos últimos no tienen pulgares oponibles, cuando un cuervo no tiene manos siquiera); por qué especies tan separadas como la nuestra y el perro muestran una sensibilidad hacia la mirada tan distinta al resto de especies (domesticación mediante, sí, pero necesito estudiar las razones); o por qué los bonobos son más sensibles al entorno social y los chimpancés al entorno físico. En definitiva, la razón por la que soy rupturista no es el antropocentrismo, sino porque no quiero observar discursos evolucionistas de pura fantasía donde la presión evolutiva que sufrieron los chimpancés hace menos tiempo que cuando nos separamos de ellos, pueden explicar nuestras propias presiones evolutivas o que si soy un padre soltero que cría a sus hijos no formo parte de un esquema donde el discurso es más importante que los matices. Menos eslabones perdidos y más patrones que nos hacen ser como somos.
Referencias:
Dunbar, R.I.M, Barrett, L. & Lycett, J. (2005). Evolutionary Psychology: A Beginner´s Guide. London: Oneworld Beginner´s Guide.
Richter, S. & Meier, R. (1994). The Development of Phylogenetic Concepts in Hennig’s Early Theoretical Publications (1947-1966). Systematic Biology, 43(2), 212–221.
Tomasello, M. (2019). Becoming Human: A Theory of Ontogeny. Harvard University Press.
Wilkins, J. S. (2009). Species: A History of the Idea. University of California Press.
Canción recomendada:

Yo te voy leyendo…. Espero que conforme lea más, entienda más y creo que me resultará más interesante incluso que lo que me puede resultar ahora🤭🤭🤣🤣…. Entiendo que yo soy muy ajeno a la materia y el nivel de los textos es elevado… De todo no se puede saber, por eso estoy «aquí». Un saludo
Me gustaMe gusta